Photobacterium profundum - Photobacterium profundum
| Photobacterium profundum | |
|---|---|
 | |
| Конфокальный изображение штамма P. profundum SS9, экспрессирующего зеленый флуоресцентный белок. Полоса: 10 мкм | |
| Научная классификация | |
| Королевство: | |
| Тип: | |
| Класс: | |
| Порядок: | Вибрионалес |
| Семья: | |
| Род: | |
| Виды: | P. profundum |
| Биномиальное имя | |
| Photobacterium profundum | |
Photobacterium profundum это глубокое море Гаммапротеобактерии, принадлежащий семье Vibrionaceae и род Фотобактерии. Как и другие представители этого рода, P. profundum является морским организмом и имеет два круговых хромосомы.[1] P. profundum это грамотрицательный стержень с возможностью роста при температуре от 0 ° C до 25 ° C и давлении от 0,1 МПа до 70 МПа в зависимости от деформации. Он нуждается в соли, способен усваивать широкий спектр простых и сложных углеводы и имеет два жгутики системы. Клетки палочковидной формы, длиной 2–4 мкм и шириной 0,8–1,0 мкм, с одним неокрашенным жгутиком.[2] Эта бактерия была первоначально выделена в 1986 г. из Сулу Море и в настоящее время существует 4 культивируемых штамма дикого типа P. profundum, (штаммы SS9, 3TCK, DJS4 и 1230).[3]
P. profundum штамм SS9 имеет оптимальный рост при 15 ° C и 28 МПа, что делает его психрофил и пьезофил. P. profundum штамм 3TCK, изоляты из Залив Сан-Диего,[4] оптимально растет при 9 ° C и 0,1 МПа и P. profundum штамм DSJ4, выделенный из Рюкю желоб от Япония на глубине 5110 м, оптимально растет при 10 ° С и 10 МПа.[2] На основе 16S рРНК последовательность P. profundum близок к роду Вибрион, наиболее заметным видом является человеческий патоген Холерный вибрион.
У штамма SS9 было показано, что несколько генов стрессовой реакции активируются в ответ на атмосферное давление, к ним относятся htpG, dnaK, dnaJ и groEL.[1] Типы и количество цепей жирных кислот в клеточной мембране также реагируют на изменения давления и температуры.[5] При низкой температуре и высоком давлении деформация SS9 увеличивает содержание моно- и полиненасыщенный жирные кислоты. Это приводит к увеличению текучести мембраны за счет уменьшения упаковки цепей жирных кислот, что приводит к образованию жидкокристаллической структуры в мембране, а не к структуре геля.[5] В белок внешней мембраны Было показано, что OmpH активируется при повышенном давлении, противоположное верно для белка внешней мембраны OmpL, который активируется в ответ на низкое давление.[6]
В 2005 году Vezzi et al. опубликовал последовательность генома для P. profundum штамм SS9. Геном P. profundum состоит из кольцевой хромосомы размером 4,1 Мбит / с, минорной кольцевой хромосомы размером 2,2 Мбит / с, а также кольцевой хромосомы размером 80 т.п. плазмида. Штамм SS9 имеет 14 генов рибосомной РНК (рРНК) на хромосоме 1 и 1 на хромосоме 2; это наибольшее количество генов рРНК, обнаруженных у любой бактерии. Хромосома 1 состоит в основном из генов, которые необходимы для роста, тогда как хромосома 2, по-видимому, представляет собой большую плазмиду, которая в эволюционном масштабе времени приобрела несколько сменные элементы.[4] В геноме P. profundum есть большое количество открытые рамки для чтения (ORF), которые уникальны для SS9 и не встречаются у других членов семейства Vibrionaceae.
Последовательность генома также выделила полный Путь Stickland для ферментации аминокислоты; это был первый раз, когда этот путь был идентифицирован в аэробный бактерия. У этой бактерии также присутствуют два полных пути АТФ-синтазы F1F0 (по одному на каждой хромосоме): это может объяснить ее способность продуцировать АТФ как при высоком, так и при низком давлении.
За этой работой в 2005 г. последовала еще одна статья Campanaro et al. который подробно микрочип работают по сравнению экспрессии генов при субоптимальных, оптимальных и сверхоптимальных температурах и давлении для штаммов SS9, 3TCK и DSJ4.[4] Campanaro et al. показали, что 544 ORF расходятся или отсутствуют в геноме 3TCK и 562 ORF расходятся или отсутствуют в хромосомах DSJ4 по сравнению с таковой у SS9. В этой статье также подчеркивается, что в 3TCK отсутствует система латеральных жгутиков, которая регулируется в SS9 при повышенном давлении, а также отсутствует 3 фаг -зависимые области от 3TCK и 4 связанные с фагом области от DSJ4.[4][7]
Транскрипционный ландшафт штамма DB110 дикого типа и штамма toxR мутант TW30 исследовали с помощью секвенирование следующего поколения.[8] ToxR - трансмембранный ДНК-связывающий белок впервые обнаружен в Холерный вибрион, где он регулирует значительное количество генов, участвующих в адаптации к окружающей среде и вирулентность. В P. profundum на количество и активность этого белка влияет гидростатическое давление, и его роль связана с регуляцией генов в зависимости от давления.[9] Результаты получены из РНК-последовательность эксперименты выявили сложный паттерн экспрессии с группой из 22 генов, имеющих профили экспрессии, подобные OmpH, который является белком внешней мембраны, транскрибируемым в ответ на высокое гидростатическое давление.[10] Более того, RNA-seq позволил глубоко охарактеризовать транскрипционный ландшафт, что привело к идентификации 460 предполагаемых малая РНК гены и обнаружение 298 генов, кодирующих белок, ранее неизвестных. Полногеномное предсказание оперон структура, сайты начала и окончания транскрипции, выявила неожиданно большое количество генов (992) с большими 5’-UTR, достаточно длинными, чтобы нести цис-регуляторный Структуры РНК, предполагающие корреляцию между размером межгенной области и длиной UTR.
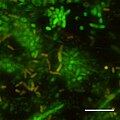
CSLM-образ начальных этапов биопленка образование в Photobacterium profundum SS9 с использованием живых / мертвых красителей. Давление = 28 МПа Температура = 15 градусов Цельсия Бар = 10 микрометров

CSLM Photobacterium profundum SS9, экспрессирующий GFP. Бар = 10 микрометров


использованная литература
- ^ а б Вецци, А. и др., Жизнь на глубине: последовательность генома Photobacterium profundum и анализ экспрессии. Science, 2005. 307 (5714): с. 1459-61.
- ^ а б Ноги Ю., Масуи Н. и Като К. Photobacterium profundum sp. nov., новый умеренно барофильный вид бактерий, выделенный из глубоководных отложений. Extremophiles, 1998. 2 (1): с. 1-7.
- ^ ДеЛонг Э.Ф. Адаптация глубоководных бактерий к глубоководной среде 1986 г.
- ^ а б c d Campanaro, S, Vezzi, A, Vitulo, N, Lauro, FM, D'Angelo, M, Simonato, F, Cestaro, A, Malacrida, G, Bertoloni, G, Valle, G, Bartlett, DH., Элементы с боковым переносом и адаптация к высокому давлению у штаммов Photobacterium profundum. BMC Genomics, 2005. 6: с. 122.
- ^ а б Аллен, Э.Е., Д. Фаччиотти и Д.Х. Бартлетт, Мононенасыщенные, но не полиненасыщенные жирные кислоты необходимы для роста глубоководной бактерии Photobacterium profundum SS9 при высоком давлении и низкой температуре. Appl Environ Microbiol, 1999. 65 (4): стр. 1710-20.
- ^ Бартлетт, Д.Х. и Т.Дж. Welch, экспрессия гена ompH регулируется множеством факторов окружающей среды в дополнение к высокому давлению в глубоководных бактериях Photobacterium разновидности штамма SS9. J. Bacteriol, 1995. 177 (4): p. 1008-16.
- ^ Campanaro, S, Vezzi, A, Vitulo, N, Lauro, FM, D'Angelo, M, Simonato, F, Cestaro, A, Malacrida, G, Bertoloni, G, Valle, G, Bartlett, DH., Элементы с боковым переносом и адаптация к высокому давлению у штаммов Photobacterium profundum. BMC Genomics, 2005. 6: с. 122.
- ^ Campanaro, S, DePascale, F, Telatin A, Schiavon R, Bartlett, DH, Valle, G. Транскрипционный ландшафт глубоководной бактерии Photobacterium profundum как у мутанта toxR, так и у его родительского штамма. BMC Genomics, 2012 13: с. 567. DOI: 10.1186 / 1471-2164-13-567.
- ^ ДиРита, VJ, Mekalanos, JJ. Периплазматическое взаимодействие между двумя мембранными регуляторными белками, ToxR и ToxS, приводит к передаче сигнала и активации транскрипции. Cell, 1991. 64 (1): p. 29–37.
- ^ Велч, Т.Дж., Бартлетт, Д.Х., Идентификация регуляторного белка, необходимого для экспрессии чувствительного к давлению гена у глубоководных бактерий Фотобактерии видовой штамм SS9. Mol Microbiol, 1998. 27 (5): с. 977–985.